Fígado e Homeostase
O fígado é o maior órgão parenquimatoso do corpo e, ao mesmo tempo, a maior glândula. Seu tamanho e complexidade se justificam pela enorme diversidade de funções que exerce, ocupando papel central na manutenção da homeostase. Esse órgão participa ativamente do metabolismo de carboidratos, lipídios e proteínas, regula a concentração de glicose no sangue, sintetiza proteínas plasmáticas essenciais, secreta bile e é responsável por processos de detoxificação. Além disso, controla o fluxo e a qualidade das substâncias absorvidas no trato digestório, funcionando como uma “barreira metabólica” antes que esses compostos alcancem a circulação sistêmica.
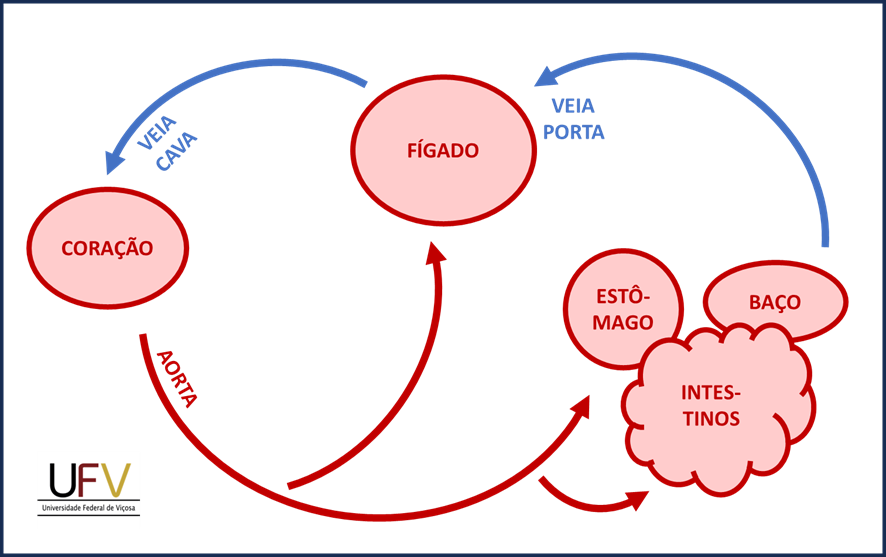
O fígado recebe sangue de duas fontes distintas: a artéria hepática, que traz sangue rico em oxigênio, e a veia porta, que conduz sangue proveniente do intestino carregado de nutrientes, metabólitos e possíveis toxinas. A veia porta constitui o chamado sistema porta-hepático. Um sistema porta no contexto da circulação sanguínea ocorre quando o sangue, após deixar o sistema arterial (de distribuição) passa por dois leitos capilares antes de ser recolhido definitivamente pelo sistema venoso (de coleta) para a veia cava. Na maioria dos órgãos do corpo, o sangue passa apenas por um leito capilar. No sistema porta-hepático, o sangue arterial deixa a aorta para o trato gastrointestinal e o baço, onde passa pelo primeiro leito capilar. Em seguida, é conduzido ao fígado pela veia porta. Os capilares hepáticos, que são do tipo sinusoide, correspondem ao segundo leito capilar. Só depois, o sangue é recolhido para a veia hepática e, daí, para a veia cava caudal.
O fígado é revestido por uma cápsula de tecido conjuntivo coberta por peritônio e a partir da qual, projetam-se fibras conjuntivas que delimitam os lóbulos hepáticos. Essa separação lobular é muito delicada na maioria das espécies, porém, em suínos, o tecido conjuntivo forma verdadeiros septos entre os lóbulos.
O lóbulo hepático é a unidade funcional do fígado e tem um formato aproximadamente hexagonal. Em seus vértices encontram-se os chamados espaços porta, que abrigam um ramo da artéria hepática, um ramo da veia porta, um dúctulo biliar e vasos linfáticos. No centro de cada lóbulo situa-se a veia centrolobular, para onde converge o sangue após atravessar os sinusoides hepáticos. Os hepatócitos organizam-se em fileiras dispostas de maneira radial, de forma que o sangue flua entre elas.
Os capilares sinusoides possuem um endotélio descontínuo e lâmina basal incompleta, permitindo uma troca intensa de substâncias entre o plasma e os hepatócitos. Ocorre mistura do sangue proveniente da arteríola portal hepática e da vênula portal hepática e ele flui do espaço porta em direção à veia centrolobular.
O espaço existente entre a parede do sinusoide e a superfície dos hepatócitos é chamado de espaço perissinusoidal (espaço de Disse)
. Nele, o plasma circula em contato próximo com as microvilosidades dos hepatócitos, favorecendo tanto a secreção quanto a absorção de substâncias. No polo oposto a este que faz contato com o plasma, a membrana celular do hepatócito fixa-se às outras células por meio de junções firmes de forma a permitir a formação de um caminho para drenagem da bile em direção ao espaço porta. Esse caminho é chamado de canalículo biliar. Lateralmente, os hepatócitos têm muitas junções firmes com as células vizinhas de forma a não permitir contato entre os sinusoides e os canalículos biliares. Microscopicamente, os hepatócitos têm núcleo redondo e centralizado, com vários pontos de heterocromatina. Hepatócitos com dois núcleos não são incomuns.

A) Lóbulo hepático; ep: espaço porta; vcl: veia centrolobular; B) Detalhe do lóbulo hepático; vcl: veia centrolobular; hc: hepatócito; cb: canalículo biliar; cs: capilar sinusoide; eD: espaço de Disse; ah: arteríola (portal) hepática; vp: vênula portal (hepática); db: dúctulo biliar; ep: espaço porta.
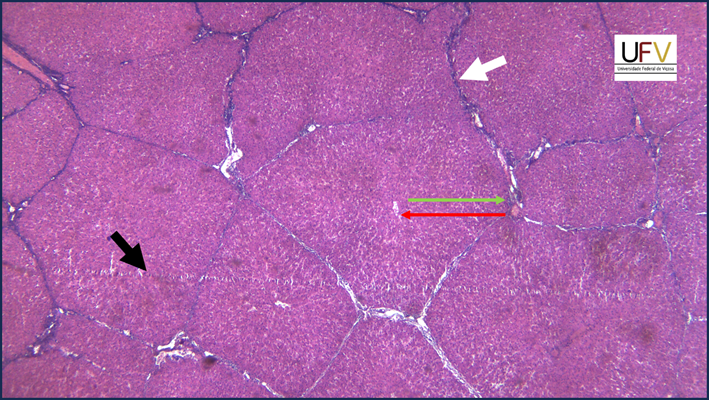
Na micrografia acima, vemos fragmento de fígado suíno, com septos conjuntivos (um deles indicado pela seta branca) delimitando os lóbulos hepáticos. A seta vermelha indica a direção em que o sangue flui, do espaço porta para a veia centrolobular. A seta verde indica o sentido que a bile percorre os canalículos, a partir dos hepatócitos perivenosos, até o dúctulo biliar. A seta preta indica uma marca horizontal que atravessa toda a imagem; esta marca é um artefato do processamento histológico, que aparece quando há um dente na navalha durante a microtomia.
Além dos hepatócitos, que constituem a maior parte do parênquima, o fígado contém outros tipos celulares importantes. As células de Kupffer são macrófagos residentes, responsáveis pela remoção de detritos e patógenos, e se encontram dentro dos capilares. As células estreladas, localizadas no espaço de Disse, armazenam vitamina A e controlam a deposição de matriz extracelular, mas podem proliferar e secretar colágeno em resposta a agressões crônicas, contribuindo para fibrose hepática. Há ainda as chamadas pit cells, um tipo de célula natural killer especializada na imunidade hepática.
Os lóbulos hepáticos podem ser divididos em zonas metabólicas de acordo com a proximidade ao espaço porta. A zona periportal, ou zona 1, mais próxima da entrada de sangue, recebe maior aporte de oxigênio e abriga hepatócitos voltados para a gliconeogênese e a síntese proteica. Nesta região estão presentes mais fibroblastos e hepatócitos menores. Células de defesa também podem ser observadas. A zona intermediária, ou zona 2, representa uma transição entre os dois polos, enquanto a zona perivenosa, ou zona 3, mais distante do suprimento arterial, contém hepatócitos com alta atividade glicolítica e abundância de enzimas do citocromo P450, sendo por isso a região mais vulnerável a hipóxia e toxinas.
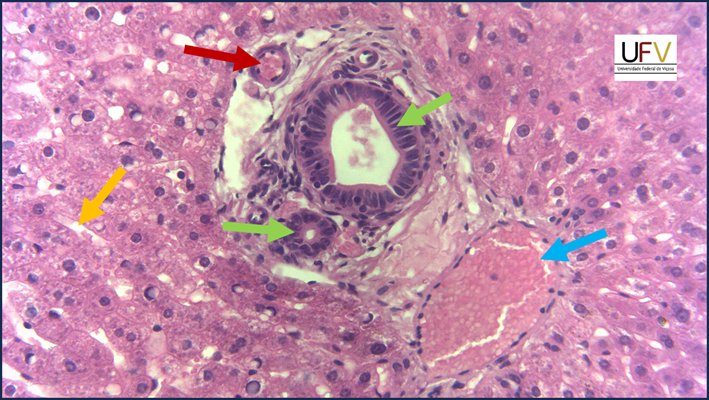
Na micrografia acima, podemos ver detalhes do espaço porta. A seta vermelha mostra uma arteríola e a azul, uma vênula – note que, em ambos, o epitélio de revestimento, ou seja, o endotélio, é pavimentoso. Os dúctulos biliares vão convergindo progressivamente e seu revestimento passa de cuboide simples para colunar simples – esses dois tipos podem ser vistos na imagem, nas setas verdes. A seta amarela mostra um capilar sinusoide, que passa entre fileiras de hepatócitos.
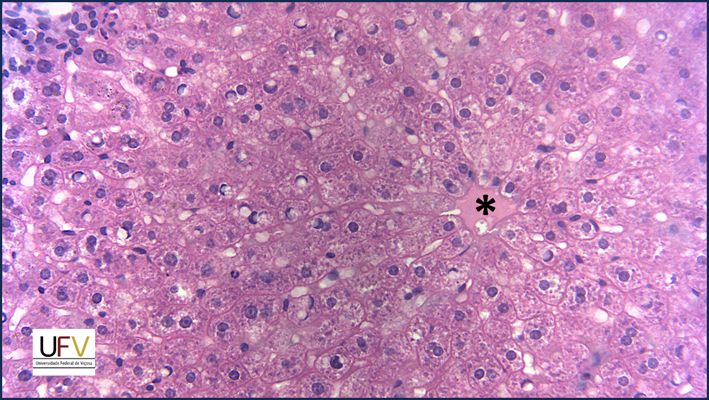
Na micrografia acima, é possível observar o padrão radial das fileiras de hepatócitos em relação à veia centrolobular. Um desafio: tente encontrar um leucócito polimorfonuclear nesse campo.
O metabolismo de carboidratos é uma das funções mais evidentes do fígado. Quando há glicose abundante no sangue, o fígado capta essa molécula por meio do transportador GLUT2 e a converte em glicogênio ou em lipídeos. Em períodos de jejum ou demanda energética aumentada, realiza a quebra do glicogênio (glicogenólise) e a gliconeogênese, produzindo glicose a partir de aminoácidos e glicerol. O transportador GLUT2 não depende de insulina para movimentar a glicose através da membrana e funciona nos dois sentidos (de fora para dentro, ou de dentro para fora) de acordo com a direção da diferença de concentração da glicose.
Quanto aos lipídeos, o fígado é capaz de sintetizar ácidos graxos, triglicerídeos e colesterol, além de embalar esses compostos em lipoproteínas para transporte sanguíneo. Lipídios de origem intestinal chegam ao fígado de forma diferente dos carboidratos e aminoácidos: por serem lipossolúveis, são incorporados a quilomícrons ainda no intestino e passam primeiro pelo sistema linfático, alcançando o fígado via circulação sistêmica, através da artéria hepática.
No metabolismo proteico, o fígado exerce funções fundamentais. Ele sintetiza diversas proteínas plasmáticas, como albumina, globulinas, fatores de coagulação e proteínas de fase aguda. Também desempenha papel no catabolismo de aminoácidos e na formação da ureia, processo essencial para a eliminação de nitrogênio em excesso.
Outra função de grande importância é a produção contínua de bile. Essa secreção contém sais biliares, fundamentais para a emulsificação das gorduras e absorção intestinal de lipídeos, além de bilirrubina e outras substâncias que o fígado precisa excretar, incluindo hormônios e xenobióticos. Em espécies que possuem vesícula biliar, como cães, a bile pode ser armazenada e liberada em resposta ao hormônio colecistoquinina durante a digestão. Outras espécies, como equinos e ratos, não possuem vesícula biliar, e a bile é liberada continuamente no intestino.
No que diz respeito à detoxificação, o fígado atua em três etapas. Na fase I, enzimas da família do citocromo P450 adicionam grupos polares (como um simples hidroxil, -OH) a moléculas lipofílicas, tornando-as mais hidrossolúveis. Esses intermediários, por vezes mais tóxicos que as moléculas originais, são então processados na fase II, em que ocorre a conjugação com açúcares ou peptídeos, tornando-os inativos e seguros. Por fim, a fase III consiste na eliminação dos compostos, frequentemente pela bile.
A atividade hepática é regulada por diferentes hormônios e pelo sistema nervoso autônomo. A insulina favorece a síntese de glicogênio e inibe a gliconeogênese e a glicogenólise. O glucagon exerce efeito oposto, estimulando a glicogenólise, a gliconeogênese e a lipólise. O hormônio do crescimento (GH) estimula a gliconeogênese, a oxidação de ácidos graxos e a síntese proteica, além de induzir os hepatócitos a produzirem o IGF-I, fundamental no eixo endócrino hipotálamo-hipófise-fígado. Os glicocorticoides também estimulam a gliconeogênese, mas utilizam aminoácidos como substrato, em detrimento da síntese proteica. Do ponto de vista neural, a inervação simpática promove glicogenólise e gliconeogênese, enquanto a parassimpática favorece o armazenamento de glicogênio.
Assim, o fígado não atua de forma isolada: sua atividade metabólica está em constante diálogo com outros órgãos, especialmente o pâncreas endócrino, que regula o armazenamento e a mobilização de glicogênio por meio da insulina e do glucagon. Além da síntese de glicose e lipídios, os hepatócitos também são responsáveis pela produção de proteínas plasmáticas fundamentais, como a albumina e os fatores de coagulação, ambos com grande relevância clínica. Uma falha na síntese de albumina leva ao desequilíbrio osmótico e ao acúmulo de líquido nos tecidos, resultando em edema generalizado (anasarca), enquanto alterações na produção de fatores de coagulação podem causar coagulopatias. Na dimensão exócrina, a secreção de bile representa a principal contribuição do fígado para a digestão, e sua interrupção compromete a absorção de lipídios e vitaminas lipossolúveis. Já no campo imunológico, as células de Kupffer destacam o fígado como um órgão linfoide secundário, atuando na defesa contra microrganismos do trato gastrointestinal; quando essa barreira falha, há risco de sepse de origem entérica. Além disso, o fígado participa da depuração de hormônios, como os estrogênios, e sua falha pode levar ao acúmulo desses compostos; em humanos, por exemplo, a cirrose está associada ao aparecimento de características sexuais secundárias atípicas. Finalmente, sua capacidade de metabolizar e neutralizar substâncias tóxicas, incluindo fármacos, posiciona o fígado como órgão central não apenas na manutenção da homeostase, mas também na interface direta com a clínica e a farmacologia veterinária.
Referências
BERG, J. M.; TYMOCZKO, J. L.; GATTO, G. J. Biochemistry. 8. ed. New York: W. H. Freeman, 2015. ISBN 978-1-118-50139-9.
DRIESSEN, A. et al. The histological structure of the liver. World Journal of Gastroenterology, v. 6, n. 1, p. 1-11, 2000. DOI: 10.3748/wjg.v6.i1.1.
HALL, J. E. Guyton and Hall Textbook of Medical Physiology. 11. ed. Philadelphia: Saunders, 2006. ISBN 0-7817-4148-3.
LISKA, D. J. The detoxification enzyme systems. Alternative Medicine Review, v. 3, n. 3, p. 187-198, 1998. ISSN 2394-5893.
MAEDA, K.; YAMAMOTO, Y.; WATANABE, M. et al. Role of liver in systemic homeostasis. Physiological Reports, v. 2, n. 9, p. 1-14, 2014. DOI: 10.1002/j.2040-4603.2014.tb00548.x.
MEIJER, A. J.; LAMMERS, W. Hepatic regulation of metabolism. Journal of Physiology, v. 598, n. 2, p. 295–310, 2020. DOI: 10.1113/JP281061.
MEYER, H. P. et al. The liver: metabolic functions and regulation. Zeitschrift für Gastroenterologie, v. 35, n. 4, p. 245–252, 1997. DOI: 10.1055/s-2007-1007204.
RAWLINGS, N. D.; SALVESEN, G. Handbook of Proteolytic Enzymes. 3. ed. Cambridge: Academic Press, 2013. DOI: 10.1016/B978-0-12-809954-4.00043-8.